



三、克隆基因表达
基因重组和克隆的最终目标通常使克隆基因在宿主细胞中高效表达,以便获得表达产物。根据受传细胞的不同,基因表达系统分为原核表达系统和真核表达系统。
1.原核表达系统
大肠杆菌表达系统是基因表达技术中发展最早、目前应用最广泛的经典表达系统。该项技术的主要方法是将已克隆入目的基因DNA段的载体(一般为质粒)转化为细菌(通常选用的是大肠杆菌),通过IPTG诱导并最终纯化获得所需的目的蛋白。外源基因能否在原核细胞中高效表达决定于原核细胞启动子强弱,拷贝数高低及核糖体结合位点等因素。以大肠杆菌lac操纵子调节机理为基础设计构建的表达系统,称为lac表达系统。其中包括lac、trp和tac启动子的作用。以λ噬菌体早期转录启动子PL、PR为核心构建的表达系统称为PL和PR表达系统。这两类启动子都具有很强的启动转录能力。比lac启动子活性强8~10倍,受λ噬菌体C1基因负调控,C1基因编码温度敏感蛋白,其类似于lac启动子的阻遏蛋白,在28~32℃时,C1基因产生正常的有功能蛋白,抑制转录进行,温度升高至42℃时,C1被破坏(可逆过程),操纵基因解除封闭,转录开始。
一般来说,细菌内基因拷贝数越多,基因表达量越高,因而可以通过提高基因拷贝数来增加表达量。如此,可以采用温度诱生性载体,这类载体在低温时拷贝数较低,温度升高后拷贝数大量增加,表达量升高。从核糖体结合位点看,翻译效率会受到某些因素影响。原核基因核糖体结合位是从转录起始位点开始延伸几十个碱基长度的一段序列。核糖体结合位点中与rRNA16亚基3′端互补的核心部分为SD序列(图4-16)。有实验表明,SD序列与起始密码子之间的距离可以显著影响mRNA的翻译效率。当lac启动子的SD顺序距AUG为7个核苷酸时表达水平最高,而间隔8个核苷酸时表达水平可降低数百倍。因此,许多大肠杆菌表达载体的启动子下游都设计了包括SD序列在内的核糖体结合位点,目标基因编码区插入SD序列下游即可实现表达。
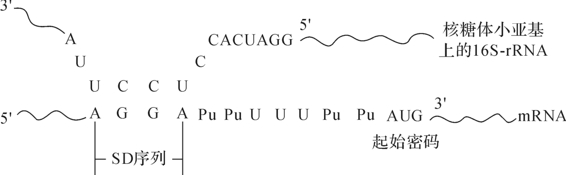
图4-16 原核生物mRNA与核糖体rRNA结合的SD序列
2.真核表达系统
原核表达系统优点在于能够在较短时间内获得基因表达产物,而且所需的成本相对比较低廉。但与此同时原核表达系统还存在许多难以克服的缺点,如通常使用的表达系统无法对表达时间及表达水平进行调控,有些基因的持续表达可能会对宿主细胞产生毒害作用,过量表达可能导致非生理反应,目的蛋白常以包涵体形式表达,导致产物纯化困难;而且原核表达系统翻译后加工修饰体系不完善,表达产物的生物活性较低。为克服上述不足,许多学者将原核基因调控系统引入真核基因调控领域,其优点是:①根据原核生物蛋白与靶DNA间作用的高度特异性设计,而靶DNA与真核基因调控序列基本无同源性,故不存在基因的非特异性激活或抑制;②能诱导基因高效表达,可达105倍,为其他系统所不及;③能严格调控基因表达,即不仅可控制基因表达的“开关”,还可人为地调控基因表达量。因此,利用真核表达系统来表达目的蛋白越来越受到重视。目前,基因工程研究中常用的真核表达系统有酵母表达系统、昆虫细胞表达系统和哺乳动物细胞表达系统。
(1)酵母表达系统
酵母是单细胞真核生物,具有真核细胞特点,可以识别内含子,对蛋白质进行多种翻译后修饰,有分泌功能。酵母又具有原核细胞培养简单、生长周期短、容易操作等特点,故是一种理想的基因工程受体细胞,也是一种很好的真核基因表达系统。已有多种蛋白质在酵母细胞中成功表达,如人乙肝病毒表面抗原、核心抗原、干扰素、人表皮生长因子和胰岛素前体等。
酵母载体中的调控序列如下:①酵母的自主复制序列(ARS),相当于复制起始区(点),可启动基因在酵母中的复制。②酵母细胞中有一种称为2μm质粒,它以高拷贝数存在于细胞核外,为小的环状DNA,长约2μm,可以独立复制与转录。常用酵母启动子有:ADH(醇脱氢酶)基因、PGK(磷酸甘油激酶)基因、GAP(3′—磷酸甘油脱氢酶)基因等,其中,DGK和GAP在细胞内各占总蛋白的5%左右,属于高含蛋白质。
酵母基因表达系统的载体通常既能在酵母菌中复制也能在大肠杆菌中进行复制,形成所谓酵母-大肠杆菌穿梭载体(shuttle vector)。穿梭载体pYE13(酵母附加体型质粒)含有整个pBR322质粒序列(图4-17)。使用pYE13构建重组时,首先转化大肠杆菌受体细胞,经筛选,鉴定和分析,再将需要的重组分子转入酵母细胞。其结构组成特点:
①DNA复制起始点:这是一段具有复制起始功能的DNA序列,来自酵母菌的2μ质粒的复印起始区及酵母基因中的自主复制序列(ARS)。(https://www.xing528.com)
②选择标记:这是筛选酵母转化子必需的构件,与宿主的基因型相互配合。酵母表达系统中所用的选择标记有两类。一类是酵母宿主为营养缺陷型,如leu2、ura3、his3、trpl等,其选择标记就是营养合成代谢途径中相应的LEU2、URA3、HIS3、TRP3。由图4-17可以看出,只有获得载体的缺陷型细胞才能在不含有氨酸的培养基中生存。另一类是星性标记,如G418和CYH,其优点是可以用野生型酵母的转化。
③整合介导区:根据不同的目的和要求,可以人为地通过特定的整合介导区序列控制载体在宿主染色体上整合的位置和拷贝数。酵母染色体的任何片段都可作为整合介导区。最方便、最常用的单拷贝整合介导区是营养缺陷型选择标记基因序列,高拷贝重复序列可作为多拷贝整合介导区。
④有丝分裂稳定区:游离载体在细胞有丝分裂时能否有效分配到小细胞中去是决定转化子稳定性的重要因素之一。有丝分裂稳定区的作用就是在细胞有线分裂时能帮助载体在子细胞之间平均分配。常用的有丝分裂稳定区是来自于酵母染色体的丝粒片段。
⑤表达盒:表达盒是酵母基因表达载体最重要的构件,主要由启动子和终止子组成。如果需要外源基因表达产物分泌,在表达盒的启动子下游还应包括分泌信号序列。由于酵母对异种生物的转录调控元件的识别和利用率很低,表达盒中的转录启动子、分泌信号序列及终止子都应该来自酵母本身。
外源基因在酵母中有两种表达形式:
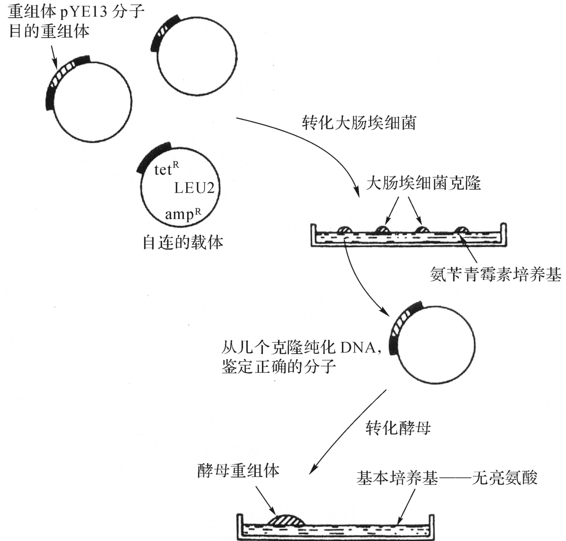
图4-17 穿梭载体DYE13d酵母中的克隆
(引自魏群等译 2003)
①直接表达外源基因:即将外源蛋白表达在细胞中,而不分泌出胞外。酵母菌表达外源基因,多为胞内表达,其中,乙肝病毒表达抗原在酵母菌中的表达就是个成功的例子。
②分泌表达:其可以避免产物被降解,同时有利于产物纯化,并帮助产物进行糖基化修饰,是一种很好的表达形式。在酵母细胞中利用外源性的信号肽很难分泌表达。然而利用酵母细胞中天然存在的分泌型蛋白质的信号肽,可以构建酵母分泌型表达载体,实现外源基因的分泌表达。最常用的酵母分泌信号肽序列是啤酒酵母α因子的信号肽。
(2)昆虫细胞表达系统
昆虫细胞表达系统(即杆状病毒表达系统)具有独特的生物学特性,日益受到人们的重视。杆状病毒只来源于无脊椎动物,虽然已发现600多种杆状病毒,但进行分子生物学研究的不到20种。杆状病毒的基因组为单一闭合环状双链DNA分子,大小为80~160kb,其基因组可在昆虫细胞核复制和转录。DNA复制后组装在杆状病毒的核衣内,后者具有较大的柔韧性,可容纳较大片段的外源DNA插入,因此是表达大片段DNA的理想载体。其中,用作外源基因表达载体的杆状病毒,目前仅限于核型多角体病毒(nuclear polyhedrosis virus,NPV)。该病毒颗粒在细胞内可由多角体蛋白包裹形成长度约1~5m的包含体病毒,呈多角体形状。核型多角体病毒有两种形式:一种为包含体病毒(occluded virus,OV),另一种则为细胞外芽生病毒(budded virus,BV)。它们在病毒感染中扮演的角色不同,包含体病毒是昆虫间水平感染的病毒形式,昆虫往往是食入污染OV的食物后引起感染。
昆虫中肠上皮局部的强碱性环境(pH=10.5),可使病毒颗粒释放蛋白酶溶解多角体。BV病毒是个体内细胞间的感染形式,由细胞芽生出BV,进入血淋巴系统中感染其他部位的细胞或直接在临近细胞内感染。
近几十年,有关杆状病毒基因结构、功能和表达调节的研究进展迅速,其中研究最深入的是苜蓿银蚊夜蛾(autogra-phacalifornica)多核型多角体病毒(multiple nuclear polyhedrosis virus,MNPV),简称AcMNPV或AcNPV。该病毒是杆状病毒科Baculoviridae的原型,是一种大的、带外壳的双链DNA病毒,能感染30多种鳞翅目昆虫,被广泛用作基因表达系统载体。其他作为表达载体的杆状病毒,主要是来自家蚕的NP~(bombyx moil,Bm-NP~)。由于家蚕幼虫体内系统适合大规模地制备生产外源蛋白,且成本低,显示出良好的应用前景。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。