



一、光合膜结构与捕光色素复合体
1.光合膜的机构与成分
(1)叶绿体的超显微结构
用10万倍电子显微镜观察叶绿体的精细结构表明,一个典型的高等植物叶绿体,其表面由一双层膜包裹着,每层膜的厚度约为2nm,具有高电子密度。两层膜之间由一厚度约为1nm,被电子光束透明的区带分隔开。在叶绿体内部,可划分为两个不同的区域结构:间质(stroma)和类囊体(thylakoid)。当人们对类囊体构成不了解之前就称作为基粒(grana)(图6-2)。间质的电子密度较小,其内含有同化CO2所需的全部酶类,是合成光合产物——碳水化合物的场所。类囊体亦是一种膜系统,即膜折叠而成,因此人们又称其为类囊体膜或光合膜。
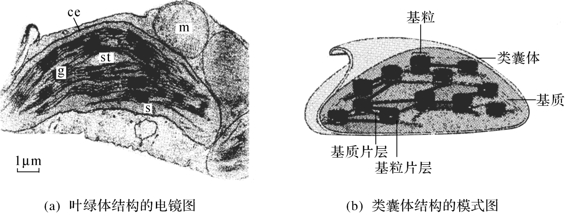
图6-2
ch:叶绿体,m:线粒体,s:基质,g:基粒,st:淀粉粒
类囊体在间质中呈不对称分布,并表现出两种不同的形态结构,一种称为基粒或基粒片层;另一种称为基质或间质膜或基质片层。基粒通常由10~100个类囊体垛叠而成,直径约为0.3~2μm。间质膜系指间质中未发生垛叠的类囊体,其上有小孔。它们贯穿在整个间质与基粒相联,形成一个三维空间的网络结构(图6-2)。
(2)类囊体的结构与成分
一般认为,类囊体由两层蛋白质分子夹着两层脂类分子构成,叶绿素呈单分子层排列在蛋白质和类脂之间,其中分布着许多电子传递体和酶类。构成类囊体的主要脂类化合物是糖脂、硫脂和磷脂,其中主要由电中性的糖MGDG和DGDG(双半乳糖基甘油二酯)组成。
类囊体膜上分布着超分子蛋白质复合体,主要包括PSⅡ反应中心、PSⅠ反中心、细胞色素b6f复合体(Cyb6/f)和ATP合成酶四大类(图6-3)。它们担负着光能吸收、传递和能量转化作用,也可说光合作用的全过程就是在光合膜和囊类体内进行的。这需得用以下更多的篇幅来展开。

图6-3 类囊体超分子蛋白复合体在膜上分布(Nield,1997)
(3)光合色素
色素也是一种脂类化合物,它们是吸收光能的基本物质。在高等植物中主要含有两大类色素:一类是叶绿素;另一类是类胡萝卜素。在藻类中除含以上两类外还有一类藻胆素。
高等植物叶绿体中含有叶绿素a和b两种。叶绿素的结构外形像个网球拍,头部由四个吡咯环和4个甲烯基组成一个卟啉环。镁原子位于吡咯的中心,尾部则连接叶醇基。卟啉环是亲水的,则叶醇基是亲脂的,两者互相垂直,这对叶绿素分子在光合膜上的有序排列非常重要。叶绿素溶于乙醇、丙酮、乙醚等有机溶剂,如果把叶绿素溶液放在光源和分光镜之间,就可看到红、橙、黄、绿、青、蓝、紫7色光谱。叶绿素有两个强的吸收光区,一个为波长640~660nm的红光部分,另一个为波波长410~470nm的蓝紫光区(图6-4(a))。
高等植物叶绿体中的类胡萝卜素有两种,即胡萝卜素和叶黄素。胡萝卜素分子式为C40H56,它有三种异构体,即α-胡萝卜素、β-胡萝卜素和γ-胡萝卜素。叶黄素是胡萝卜素的衍生物,其分子式为C40H56O2,它们在440nm和480nm间有两个光谱吸收峰(图6-4(b))。类胡萝卜素所吸收的光能传递给叶绿素用于光合作用并具有保护叶绿素的光破坏作用。
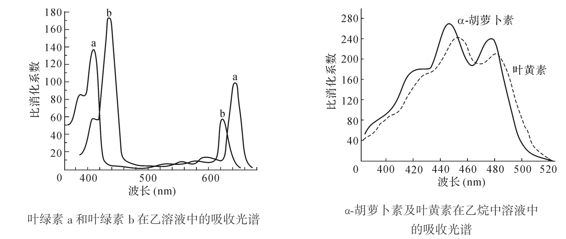
图6-4
藻胆蛋白亦称藻胆素(phycobilins),它存在于蓝藻、红藻、隐藻和一些原绿球藻中,主要有4类:藻红蛋白(PE)、藻红蓝蛋白(PEC)、藻蓝蛋白(PC)和别藻蓝蛋白(APC)。藻胆蛋白的颜色是由它们的发色团决定的,发色团是开链四吡咯化合物,称为藻胆素。藻胆蛋白主要吸收波长为450~660nm的光,其中PE的吸收峰在490~570nm,PC在620nm和APC在680nm各有一个主吸收峰。(https://www.xing528.com)
2.捕光色素复合体
捕光色素(light harvesting pigment)又名天线色素(antenna pigment),起捕获光能作用的色素和蛋白质形成的复合体,被称为捕光色素复合体,简称LHC。光系统PSⅡ和PSⅠ各自含有LHCⅡ和LHCⅠ。
(1)LHCⅡ
在叶绿素和类胡萝卜素蛋白复合体中,一个多肽链一般结合几个叶绿素分子和一个或几个类胡萝卜素分子。通常认为LHCⅡ主要包含4个组分:LHCⅡa、LHCⅡb、LHCⅡc、LHCⅡd。其中LHCbⅡ被称为大量(主体)LHCⅡ,而其余被称为微量LHCⅡ。根据编码LHCⅡ蛋白质组分的基因类型对LHCⅡ的多肽组分进行了比较系统的命名,并认为LHCⅡ主要由6种色素蛋白复合体组成,分别是Lhcb1、Lhcb2、Lhcb3、Lhcb4、Lhcb5、Lhcb6,所以,Lhcb系列为PSⅡ的LHC蛋白。
在捕光色素家族中,Lhcb1是含量最多的成员,Kuhlbrandt等(1994)已成功地对Lhcb1的结构进行了近原子分辨率(0.34nm)的解析,并提出一个较通用的结构模型,因为它们都是结构相似的一类蛋白质。初步确认其他的脱辅基蛋白含232个氨基酸残基,表观分子量约为28KD,具有3个跨膜α螺旋,80%组分为多肽,含12个叶绿素(7个chla、5个chlb)和2个叶黄素分子。其色素分子位置已基本确定,两个叶黄素分子位于中心,周围围绕着12个chl分子,所有的chlb分子均与chla分子相邻,其卟啉环之间的最短距离为0.40~0.50nm,而Mg中心之间的距离为0.83~1.05nm。
通常从植物中提取的Lhcb1以三聚体形式存在,包括Lhcb1、Lhcb2和少量Lhcb3,三种分子量分别在28KD、27KD和25KD的多肽亚基,并以随机的方式在体内组成异质三聚体。微量LHCⅡ包括Lhcb4、Lhcb5和Lhc6,在结构上是距离反应中心较近的天线系统称内周捕光色素蛋白复合体(CP43和CP47),而主体捕光色素蛋白复合体则距离反应中心较远,称为外周天线色素复合体(LHCⅡ)。因此,微量捕光复合物还起着连接主体天线色素蛋白与PSⅡ反应中心色素蛋白,将外周天线复合体捕获的光能传为PSⅡ反应中心的作用。
目前,关于LCHⅡ的结构表达方式可能有多种类型。MalKin和Niyogi(2000)提出的绿藻和高等植物所共同的一种简易图解(图6-5)。在PSⅡ中,通过光谱和生化计算大约含有单体结合的220~230个捕光色素分子作为一个光合单位。PSⅡ核心天线含有40~50个叶绿素分子,在CP43/CP47的周围结合有等量的Lhcb3、Lhcb4、Lhcb5和Lhcb6蛋白,并形成三聚体。近些年来,我国学者匡廷云等人(2003)年在光系统捕光色素蛋白体结构方面也做了不少工作,并报道了菠菜光合体系-Ⅱ(LHCⅡ)2.72A°高分辨的晶体结构。该结构显示,每个LHCⅡ单体中的14个叶绿素为8个chla和6个chlb分子,而复合物中每个色素的位置(含三聚体的类胡萝卜素)已被准确绘制成图(刘俊峰2004)。
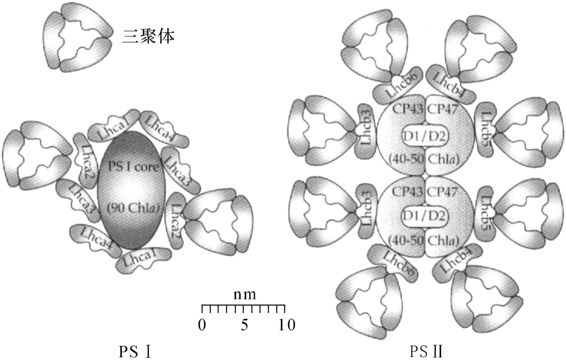
图6-5 LHCⅠ和LHCⅡ的结构
(Malkin&Niyogi,2000)
(2)LHCⅠ
1980年Muler等首次从高等植物中分离出了PSⅠ和LHCⅠ。LHCⅠ由几种多肽组成,结合有叶绿素和类胡萝卜,其chla和chlb两者比例约为4:1。LHCⅠ蛋白均由核基因编码蛋白在细胞质内合成后运送进入叶绿体内。从基因序列分析结果看,这些基因属捕光色素蛋白CAB基因家族。因此,LHCⅠ与LHCⅡ属于同一类蛋白。LHCⅠ同LHCⅡ有30%到40%的相同序列,局部地区,则高达60%。匡廷云等的实验结果表明,LHC1多肽组成的分子量为21~24KD,含chl21KD的色素多肽是PSⅠ长波荧光(730nm)发射的来源,而含ch1的23KD、24KD蛋白则是PSⅠ短波长荧光(680nm)发射的来源。
根据图6-5结构,Lhca系列为PSⅠ和LHC蛋白,它们与Chla、Chlb和类胡萝卜结合,形成捕光色素蛋白复合体(LHCⅠ)。也就是说在PSⅠ中,Lhca1和Lhca4结合在一起,形成LHCI-730;Lhca2和Lhca3结合在一起,形成LHCI-680。两者的77K荧光光谱分别在730nm和680nm有一最大收吸峰。电镜研究表明每个PSⅠ核心复合体可能结合有8个LHC蛋白。PSⅠ核心天线含有80~100个叶绿素分子,外周被4组Lhca蛋白包围,Lhca2蛋白可能结合Lhcb、Lhcb2成三聚体。
(3)藻类LHC
藻类的LHC种类较多,一般可分为两类:叶绿素/类胡萝卜素蛋白复合体(多数藻类)和藻胆体(红藻和蓝藻)。多数藻类LHC结构尚未清楚,只有绿藻由于其LHC与高等植物相比有很大的同源性,因此,它们在结构上也具有相似性。
叶绿素a/b—类胡萝卜素蛋白复合体主要分布在绿藻门、裸藻门和塔胞藻纲中。绿藻LHC和高等植物LHC的差别主要表现在色素组成上,如有的海洋绿藻含有管藻黄素和管藻素,而高等植物没有。裸藻的LHC蛋白与绿藻非常相似,同时含有LHC1和LHCⅡ,但两类色素的组成不大相同。叶绿素a/c—类胡萝卜素蛋白复合体主要分布在褐藻门、隐藻门、硅藻纲、金藻纲、黄藻纲等杂色藻中。红藻的主要捕光色素蛋白复合体是藻胆体。
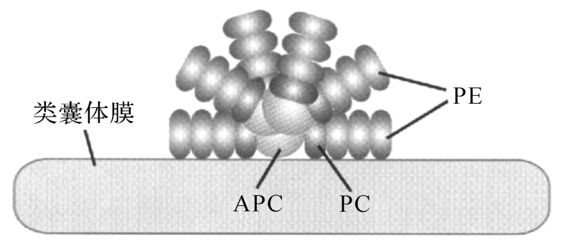
图6-6 藻胆体的结构
藻胆体是一群结合在类囊体外颗粒状的藻胆蛋白聚集物。每一个藻胆体中则含有几种不同的藻胆蛋白。蓝藻和红藻的藻胆体均含有APC,同时还含有C-PC或R-PC,部分蓝藻和多数红藻的藻胆体中还含有PE(或PEC)。此外,藻胆体中还含有无色蛋白,能够将藻胆蛋白连接起来,叫连接多肽。所以,藻胆体是由藻胆蛋白和连接多肽共同组成,分别占藻胆体的85%和15%,光能被胆藻体捕获后要传递到反应中心,其传递顺序是:PE(或PEC)→PC→APC叶绿素(图6-6)。
免责声明:以上内容源自网络,版权归原作者所有,如有侵犯您的原创版权请告知,我们将尽快删除相关内容。